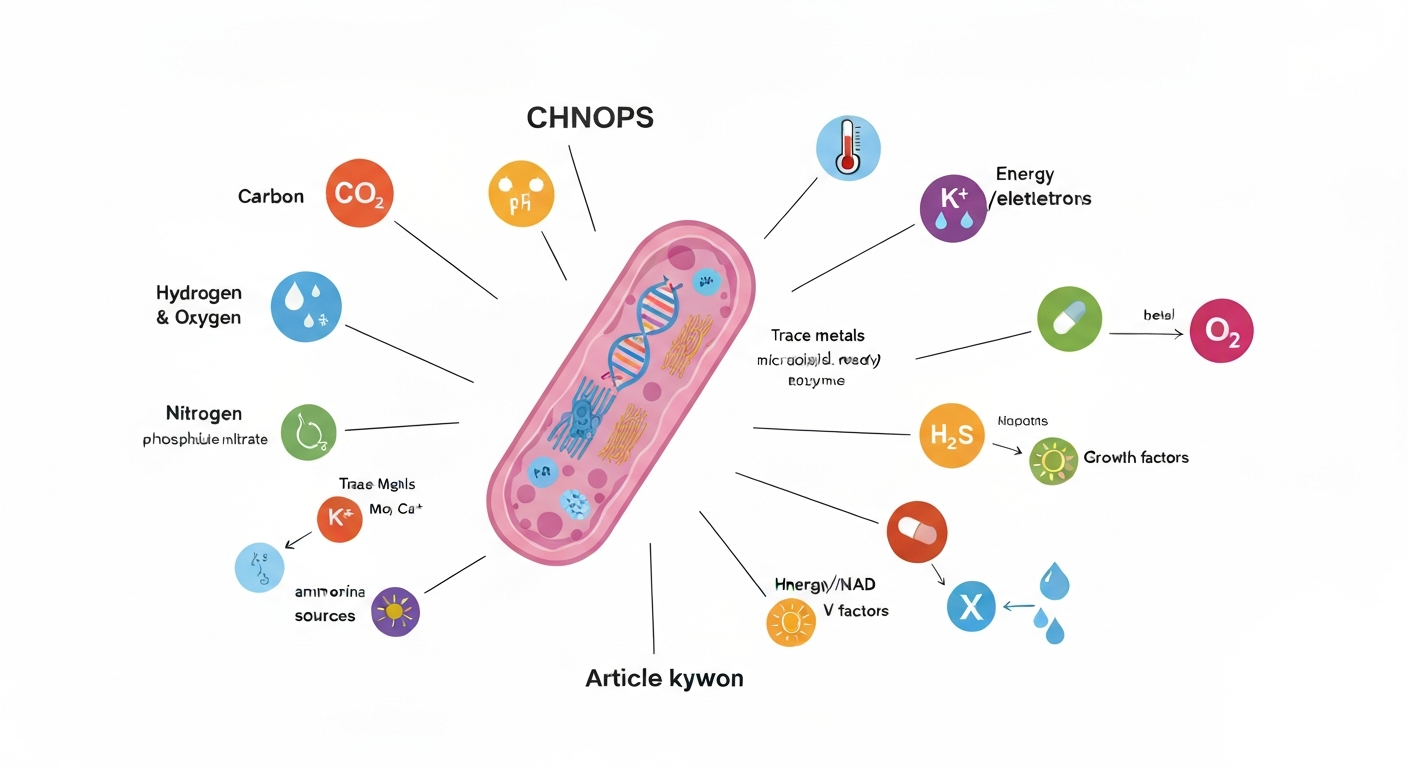
Bacteria need six core macronutrients to grow: carbon, hydrogen, oxygen, nitrogen, phosphorus, and sulfur (often abbreviated CHNOPS). Beyond those, they require trace metals like iron, zinc, and manganese, a usable energy source, and in many cases specific growth factors such as vitamins or amino acids they cannot synthesize themselves. Without all of these ingredients in place, bacterial cells stall. Understanding exactly what they need, and why, is one of the most useful frameworks in microbiology, whether you are studying for an exam, teaching a lesson, or trying to understand why food goes bad.
what nutrients do bacteria need to grow: essential guide overview

Marcus Holloway
15 Jul 2026
Who this guide is for and what you will get from it
This article is written for students, educators, and anyone who has ever wondered why bacteria thrive on a warm, protein-rich surface but not on a dry, sterile countertop. You will find clear definitions of every major nutrient category, a labeled reference table, worked examples using real species (including foodborne pathogens), and a section on how environmental conditions like temperature and pH interact with those nutrient needs. If you have already read about whether bacteria grow better in light or dark, or what food-poisoning bacteria specifically require to multiply, this article sits one level deeper, explaining the underlying biology that makes all of those questions make sense.
The four pillars of bacterial growth at a glance
Before going element by element, it helps to see the big picture. Microbiologists typically organize bacterial requirements into four overlapping categories. None of these categories works in isolation; a cell with plenty of carbon but no iron will still fail to build essential enzymes.
- Nutrients: the raw materials for building cellular structures and running metabolism (CHNOPS, trace metals, vitamins).
- Energy and electron sources: the fuel that drives biosynthesis, whether that fuel is an organic molecule like glucose, an inorganic compound like ammonia, or light.
- Physical environment: the temperature, pH, moisture level, and oxygen availability that determine whether enzymes function and membranes stay intact.
- Growth factors: specific organic compounds (vitamins, amino acids, nucleotides) that some bacteria cannot manufacture themselves and must absorb from their surroundings.
Think of it like cooking. You need ingredients (nutrients), a heat source (energy), the right kitchen conditions (physical environment), and certain pre-made components you cannot easily make from scratch (growth factors). Remove any one of these, and the dish does not come together. Bacteria are no different.
The essential macronutrients: CHNOPS explained
These six elements make up the vast majority of bacterial dry mass. Every protein, nucleic acid, lipid, and carbohydrate in a bacterial cell is built from some combination of them. Here is what each one does and where bacteria typically obtain it.
Carbon
Carbon is the structural backbone of virtually every organic molecule in the cell. Heterotrophic bacteria (which include most species you will encounter in food safety or clinical contexts) pull carbon from organic sources: sugars like glucose, amino acids, organic acids, or more complex molecules like proteins and fats. Autotrophic bacteria, by contrast, fix carbon directly from carbon dioxide (CO2), building organic compounds from scratch. The distinction matters: a heterotroph such as Escherichia coli needs a ready supply of organic carbon, while a specialist like Thiobacillus can grow on nothing but inorganic compounds and air.
Hydrogen and oxygen
These two are usually supplied together through water and dissolved oxygen, though their roles differ. Hydrogen is incorporated into organic molecules and is central to the redox reactions that generate energy. Oxygen is needed both as a building block for biomolecules (sugars, amino acids, and nucleotides all contain oxygen atoms) and, in aerobic bacteria, as the final electron acceptor in respiration. Anaerobic bacteria avoid or even tolerate oxygen poorly; they use alternative electron acceptors such as nitrate, sulfate, or carbon dioxide instead.
Nitrogen
Nitrogen is essential for amino acids (and therefore all proteins) and for nucleotides (the building blocks of DNA and RNA). Most bacteria satisfy their nitrogen requirement from ammonium ions (NH4+) or nitrate from the environment. Some bacteria, like certain Azotobacter and Rhizobium species, are nitrogen fixers: they can pull N2 directly from the atmosphere using the nitrogenase enzyme, an energetically expensive but remarkable feat. In defined laboratory media such as M9 minimal medium, NH4+ is the standard nitrogen source.
Phosphorus
Phosphorus is the backbone of the phospholipid membrane and of ATP (adenosine triphosphate), the cell's primary energy currency. It is also present in every nucleotide in DNA and RNA. Bacteria absorb it almost exclusively as inorganic phosphate (PO43-), which is why phosphate salts appear in virtually every bacteriological growth medium. In natural environments, phosphorus is often a limiting nutrient, meaning its scarcity caps how much a bacterial population can grow even when carbon and nitrogen are plentiful.
Sulfur
Sulfur is required for two of the twenty standard amino acids (cysteine and methionine) and for several enzyme cofactors. Most bacteria obtain it from inorganic sulfate (SO42-) or from organic sulfur-containing compounds. A subset of bacteria, including sulfate-reducing species like Desulfovibrio, use sulfate as a terminal electron acceptor during respiration, producing hydrogen sulfide (H2S) as a byproduct. That distinctive rotten-egg smell in waterlogged soil or improperly stored food is a direct biological signal of sulfur-based anaerobic metabolism.
Bulk cations: the often-forgotten secondary elements
Standard microbiology texts like Brock's Biology of Microorganisms distinguish a second tier of inorganic requirements beyond CHNOPS: the bulk cations potassium (K+), magnesium (Mg2+), calcium (Ca2+), and sodium (Na+). These are needed in millimolar quantities, placing them between macronutrients and trace elements. Magnesium is especially important because it stabilizes ribosomes and acts as a cofactor for enzymes involved in phosphate transfer, including those needed for DNA replication. Potassium helps maintain osmotic balance across the cell membrane. Calcium supports spore formation in species like Bacillus and plays structural roles in the cell envelope of gram-negative organisms. Sodium is essential for halophiles (salt-lovers) and plays roles in membrane transport across many species.
Trace metals and vitamins: small amounts, enormous consequences
Trace elements are needed only at microgram-per-liter to nanomolar concentrations, but remove them and bacterial growth halts entirely. The most commonly required trace metals are iron (Fe), manganese (Mn), zinc (Zn), copper (Cu), cobalt (Co), molybdenum (Mo), nickel (Ni), and sometimes selenium (Se). Each one functions as a cofactor embedded in specific enzymes, proteins, or electron carriers.
Iron deserves special attention. It is required by virtually all bacteria for cytochromes (proteins central to electron transport and energy generation) and for catalase (an enzyme that breaks down the toxic molecule hydrogen peroxide). Because free iron is scarce in both human tissues and natural environments, bacteria have evolved remarkably sophisticated uptake systems. They secrete small molecules called siderophores that scavenge iron from the environment and haul it back inside using specialized outer-membrane receptors called TonB-dependent transporters. This iron competition is one reason your immune system deliberately sequesters iron during infection: starving a pathogen of iron is a front-line defense.
Zinc is embedded in dozens of metalloenzymes involved in gene regulation and DNA repair. Manganese plays a role in oxidative stress defense. Molybdenum is the metal center of nitrogenase (for nitrogen fixation) and of nitrate reductase. Cobalt is at the heart of cobalamin (vitamin B12), and bacteria that cannot synthesize B12 must import it. Nickel is required by urease, an enzyme used by stomach colonizers like Helicobacter pylori to neutralize stomach acid. In defined laboratory media, a trace mineral mix (such as Wolfe's mineral solution) provides all of these elements simultaneously at micromolar to nanomolar effective concentrations.
B vitamins are also required by many bacteria, though the specific vitamins vary by species. Thiamine (B1), biotin (B7), riboflavin (B2), and pantothenate are commonly needed for coenzyme function. Bacteria that can synthesize these vitamins themselves are called prototrophs; those that cannot and must import them are called auxotrophs for that vitamin. The term auxotrophy simply means a nutritional dependency, and it applies to vitamins, amino acids, nucleotides, or any other compound a cell cannot make.
Energy and electron sources: how bacteria fuel themselves
Nutrients provide the raw materials, but energy drives the construction. Bacteria are classified by where they get their energy and which molecules donate the electrons that power metabolism. These two axes together determine a species' nutritional lifestyle.
Phototrophs vs. chemotrophs
Phototrophs harvest light energy, converting it into chemical energy (ATP). Familiar examples include cyanobacteria, which perform oxygenic photosynthesis and were largely responsible for oxygenating Earth's early atmosphere. Chemotrophs, which include the majority of medically and environmentally important bacteria, obtain energy by oxidizing chemical compounds. Most bacteria you encounter in food safety, clinical microbiology, or classroom experiments are chemotrophs.
Organotrophs vs. lithotrophs
Within chemotrophy, the electron donor (the compound being oxidized to release energy) may be organic or inorganic. Chemoorganotrophs, also called heterotrophs, oxidize organic molecules like glucose, amino acids, or fatty acids. This group includes E. coli, Salmonella, Staphylococcus aureus, and virtually all the pathogens relevant to food safety. Chemolithotrophs (or chemoautotrophs) oxidize inorganic compounds. Nitrosomonas europaea oxidizes ammonia (NH3) to nitrite as its energy source. Thiobacillus oxidizes sulfur compounds. Iron-oxidizing bacteria cycle between Fe2+ and Fe3+. These organisms are environmentally important in nutrient cycling but are not typically the bacteria students encounter in food-safety scenarios.
Respiration vs. fermentation
Bacteria also differ in how they dispose of electrons at the end of metabolism. Aerobic respiration passes electrons to molecular oxygen (O2), yielding the most ATP per glucose molecule. When oxygen is absent or limiting, bacteria with the right enzymes can switch to anaerobic respiration using alternative electron acceptors: nitrate (NO3-) is used first because it offers the highest energy yield, followed by manganese(IV) and iron(III) oxides, then sulfate (SO42-), and finally CO2 (a process that leads to methane production in methanogens, which are technically archaea rather than bacteria). Fermentation is different again: there is no external electron acceptor at all. The cell regenerates its electron carriers internally by converting pyruvate into lactic acid, ethanol, or other organic byproducts. Yogurt, sauerkraut, and many other fermented foods exist because of lactic acid fermentation by bacteria like Lactobacillus.
Growth factors and auxotrophy: when bacteria cannot make their own building blocks
Some bacteria are nutritionally self-sufficient prototrophs: given the right inorganic ions and a carbon source, they can synthesize every amino acid, vitamin, and nucleotide they need. Others are auxotrophs, meaning they have lost (or never evolved) the ability to make one or more essential compounds and must import them from their environment. Auxotrophy is not a flaw; in nutrient-rich niches like the human body or rotting food, it is simply more efficient to absorb a ready-made vitamin than to spend ATP synthesizing it.
The clinical textbook example is Haemophilus influenzae, which requires two specific growth factors labeled X and V. The X factor is hemin (a form of heme, required for making cytochromes), and the V factor is NAD (nicotinamide adenine dinucleotide, an essential electron carrier). H. influenzae cannot grow on ordinary blood agar because it cannot access the heme locked inside intact red blood cells; it grows readily on heated blood agar (chocolate agar) where the cells are lysed and both factors are released. This is directly relevant to clinical microbiology: identifying whether an isolate needs X, V, or both is a key step in identifying Haemophilus species.
Other common growth-factor dependencies include: requirements for specific amino acids (many Lactobacillus species need several pre-formed amino acids and are used industrially precisely because those requirements are so precise), requirements for purines or pyrimidines (nucleotide bases for DNA and RNA), and requirements for lipoic acid or cobalamin. In food safety, it is worth noting that certain pathogens thrive in protein-rich environments partly because those environments supply ready-made amino acids the bacteria would otherwise have to synthesize.
Nutrient reference table: what bacteria need and why
| Nutrient / Element | Category | Typical Source | Key Biochemical Role |
|---|---|---|---|
| Carbon (C) | Macronutrient | Glucose, organic acids, CO2 (autotrophs) | Backbone of all organic biomolecules; energy source in heterotrophs |
| Hydrogen (H) | Macronutrient | Water, organic compounds | Incorporated into organic molecules; proton gradients drive ATP synthesis |
| Oxygen (O) | Macronutrient | Water, dissolved O2, organic compounds | Component of biomolecules; terminal electron acceptor in aerobic respiration |
| Nitrogen (N) | Macronutrient | NH4+, NO3-, N2 (nitrogen fixers) | Required for amino acids, nucleotides, and cell wall components |
| Phosphorus (P) | Macronutrient | Inorganic phosphate (PO43-) | Backbone of DNA/RNA; phospholipid membranes; ATP energy currency |
| Sulfur (S) | Macronutrient | Sulfate (SO42-), organic S compounds | Component of cysteine, methionine, and enzyme cofactors |
| Potassium (K+) | Bulk cation | Potassium salts in medium/environment | Osmotic regulation; enzyme activation |
| Magnesium (Mg2+) | Bulk cation | Magnesium salts | Ribosome stability; cofactor for phosphate-transfer enzymes |
| Calcium (Ca2+) | Bulk cation | Calcium salts | Spore formation (Bacillus); cell envelope stability |
| Iron (Fe) | Trace metal | Ferrous/ferric salts; siderophore-chelated iron | Cytochromes; catalase; electron transport; oxygen sensing |
| Manganese (Mn) | Trace metal | Manganese salts (μg/L range) | Oxidative stress defense (Mn-SOD); enzyme cofactor |
| Zinc (Zn) | Trace metal | Zinc salts (μg/L range) | Metalloenzymes; gene regulation; DNA repair |
| Copper (Cu) | Trace metal | Copper salts (μg/L range) | Oxidase enzymes; electron transport in some species |
| Cobalt (Co) | Trace metal | Cobalt salts; cobalamin (B12) | Core of vitamin B12; coenzyme B12-dependent reactions |
| Molybdenum (Mo) | Trace metal | Molybdate salts | Nitrogenase (N2 fixation); nitrate reductase |
| Nickel (Ni) | Trace metal | Nickel salts (μg/L range) | Urease; hydrogenase enzymes |
| Thiamine (B1) | Vitamin / Growth factor | Synthesized or imported | Cofactor for carbohydrate metabolism enzymes |
| Biotin (B7) | Vitamin / Growth factor | Synthesized or imported | Cofactor for carboxylation reactions (fatty acid synthesis) |
| Cobalamin (B12) | Vitamin / Growth factor | Imported (many species cannot synthesize) | One-carbon metabolism; DNA synthesis |
| Hemin / X factor | Growth factor (auxotrophic) | Lysed blood cells; chocolate agar | Precursor for cytochromes; required by Haemophilus spp. |
| NAD / V factor | Growth factor (auxotrophic) | Host tissues; enriched media | Central electron carrier; required by Haemophilus spp. |
How environment and nutrients work together
A common misconception is that nutrients and environmental conditions are separate checklists. In reality, the physical environment determines whether nutrient uptake and metabolism are even possible. A bacterium surrounded by nitrogen and carbon sources will still not grow if the temperature is wrong, because the enzymes responsible for importing and processing those nutrients are proteins, and proteins denature outside their working range. Here is how each major environmental factor connects to nutrient biology.
Temperature
Temperature controls enzyme kinetics and membrane fluidity. Bacteria are broadly classified as psychrophiles (optima at or below 15°C), mesophiles (optima around 20 to 45°C), thermophiles (optima above 45°C, up to around 80°C), and hyperthermophiles (optima above 80°C). E. coli, Salmonella, and most food-poisoning bacteria are mesophiles with optima close to 37°C (human body temperature), which is why refrigeration (typically 1 to 4°C) slows their growth so effectively. ATCC product / culture growth conditions (Escherichia coli example) report Escherichia coli growth optima at approximately 37°C. Listeria monocytogenes is a dangerous exception with a reported growth range of approximately 1 to 45°C, meaning it can still import nutrients, run its metabolism, and multiply at refrigerator temperatures, just more slowly. Whether an incubator is needed for classroom bacterial growth largely depends on which species you are working with and whether room temperature falls within its optimal range.
pH
Most bacteria are neutralophiles, growing best between pH 6 and pH 8. Nutrient transporters, outer-membrane receptors, and metabolic enzymes are all pH-sensitive. Acidic or alkaline conditions can denature transport proteins directly, even if all nutrients are present. Acidophiles (optimum pH below 3) and alkaliphiles (optimum above pH 9 to 10) maintain their cytoplasmic pH within a narrower, functional range through active ion pumping, effectively shielding their internal chemistry from extreme external conditions. This is why lowering the pH of food through fermentation or adding acid (as in pickling) is such an effective preservation strategy: it does not necessarily destroy bacteria instantly, but it cuts off nutrient metabolism and growth.
Moisture and water activity
Water is not just a solvent for nutrients; it is a nutrient itself (providing H and O) and is the medium through which virtually all other nutrients are transported across the cell membrane. Bacteria require dissolved nutrients, meaning nutrient uptake simply cannot happen in dry conditions. This is why drying or salting food reduces bacterial growth: these processes reduce the water activity (aw), making less water available for microbial metabolism. Most pathogenic bacteria require an aw above roughly 0.91. When you see salt or sugar used as preservatives, they are working partly by competing for water molecules and making them unavailable to bacterial cells.
Oxygen availability
As covered in the respiration section above, oxygen availability determines which metabolic pathways are open to a bacterium. For aerobes, restricted oxygen means restricted energy production, which indirectly limits the cell's ability to run nutrient import systems (many of which are powered by ATP). Anaerobes switch to alternative electron acceptors and may actually be harmed by oxygen because they lack the enzymes to neutralize reactive oxygen species. Facultative anaerobes like E. coli shift their metabolism in response to oxygen availability, adjusting enzyme production and nutrient use accordingly. Because light affects oxygen levels in some environments (for example through photosynthesis in ponds), light exposure can indirectly influence which bacterial species are competitive in a given niche.
How bacterial nutrient needs compare to fungi and protoctists
Bacteria, fungi, and protoctists all require CHNOPS, trace metals, energy, and in many cases specific growth factors. The key differences lie in scale and complexity. Fungi are eukaryotes: their cells are larger, structurally more complex, and their nutrient requirements reflect that complexity. Many fungi require sterols (like ergosterol) for their cell membranes, a requirement bacteria do not share. Fungi are exclusively heterotrophs and typically grow more slowly than bacteria, making them better competitors in nutrient-poor environments where slow, steady absorption pays off. Protoctists (single-celled eukaryotes including algae, amoebae, and related organisms) show enormous nutritional diversity, ranging from photosynthetic algae that need little beyond light, CO2, and mineral salts, to predatory amoebae that engulf bacteria as their nutrient source. The core nutritional logic, however, is the same across all three: carbon for structure, nitrogen for proteins, phosphorus for energy transfer, trace metals for enzyme function, and appropriate environmental conditions to make it all work. For another relevant comparison, see do you need an incubator to grow bacteria.
Practical takeaways for food safety and the classroom
Understanding what bacteria need to grow turns food safety from a list of rules into a set of principles you can reason about. Most foodborne pathogens are mesophilic chemoorganotrophs: they thrive between 5°C and 60°C (the danger zone), at near-neutral pH, in moist, nutrient-rich conditions. For a focused summary of the specific conditions and controls relevant to foodborne pathogens, see our guide on what does food poisoning bacteria need to grow. Removing or limiting any one of those conditions reduces growth. Cooking kills bacteria by denaturing proteins. Refrigeration slows enzyme kinetics and nutrient uptake. Acidification (vinegar, fermentation) disrupts pH-sensitive transporters. Drying or adding salt reduces water activity and blocks dissolved nutrient access. Vacuum packaging removes oxygen and forces aerobic pathogens to switch to less efficient anaerobic metabolism, or halts growth entirely. Each of these strategies targets one or more of the fundamental nutrient and environmental requirements described throughout this article. For classroom experiments, the same logic applies: providing or withholding specific nutrients, adjusting temperature, or altering pH lets students directly observe how each condition affects growth, making abstract biochemistry visible on an agar plate.
FAQ
What are the core macronutrients bacteria need to build cells?
Bacterial biomass requires carbon, hydrogen, oxygen, nitrogen, phosphorus and sulfur (CHNOPS). Carbon provides the backbone for organic molecules; nitrogen for amino acids and nucleotides; phosphorus for nucleic acids, ATP and membranes (phospholipids); sulfur for some amino acids and cofactors; hydrogen and oxygen are in nearly all cellular compounds and in respiratory chemistry. These are the basis of minimal defined media used in microbiology.
Which inorganic ions and trace elements are essential and why?
Common bulk cations include potassium, magnesium, calcium and sometimes sodium; they stabilise ribosomes, membranes and enzymes. Essential trace metals include iron, manganese, zinc, copper, cobalt, molybdenum, nickel and occasionally selenium — they act as enzyme cofactors, redox centre components and in stress defenses. Cells use high‑affinity uptake systems (e.g., siderophores, ABC transporters) to acquire scarce metals.
Do bacteria need vitamins and other organic growth factors?
Many bacteria synthesise required vitamins and cofactors, but some are auxotrophic and must obtain specific vitamins, amino acids or nucleotides from the environment. Fastidious clinical species (e.g., Haemophilus) can require haem (X factor) and/or NAD (V factor). Growth‑factor requirements are used diagnostically and in media formulation.
How do bacteria obtain energy and electrons?
Bacteria use diverse metabolic modes: heterotrophy (organic compounds supply carbon and electrons), chemolithotrophy (inorganic compounds such as H2, Fe2+, NH3, reduced sulfur supply electrons), phototrophy (light-driven energy capture), and combinations (photoheterotrophy, chemoorganoheterotrophy). The electron acceptor used for respiration (O2, NO3−, Mn/Fe oxides, SO4 2−, CO2) depends on redox availability and determines energy yield.
How do environmental conditions interact with nutrient requirements?
Temperature affects enzyme kinetics and membrane fluidity and therefore rates of nutrient uptake and metabolism (categories: psychrophiles, mesophiles, thermophiles). pH influences nutrient solubility and cytoplasmic pH homeostasis; moisture is required for solute diffusion and metabolism; oxygen availability determines whether aerobic respiration (high energy yield) or anaerobic pathways are used; light is relevant to phototrophs. Nutrient forms (e.g., organic vs inorganic nitrogen) and availability are shaped by these conditions.
What concentrations of nutrients do bacteria typically need in defined media?
Defined minimal media (e.g., M9) supply macronutrients in millimolar ranges (carbon sources like 10–20 mM glucose), salts at millimolar levels (Mg2+, Ca2+), and phosphates at low millimolar. Trace metals and vitamins are effective at micromolar to nanomolar concentrations (Fe, Mn, Zn, Cu, Mo, etc.). Exact amounts vary by species and metabolic state.


